| 寄居蟹資訊網 | 台灣博物學家 |
動物園雜誌 42:4-14 (1991)
Shih, H.-T., 1991. Utilization of shell resource by hermit crabs in Kenting, Taiwan. Taipei Zoo Quarterly 42: 4-14.
惱人的 “房事” ─ 墾丁地區寄居蟹殼資源之利用
施習德
中山大學海洋生物研究所
不論是童年的回憶或者是前陣子炒得很熱鬧的無殼蝸牛,相信你對寄居蟹這種小動物必然不會陌生。牠總是背著牠 “寄居” 的貝殼生活,遇到敵人就立刻縮入殼中,等周圍沒有動靜了,才偷偷摸摸探出頭來悄悄的離去。這樣子似乎太懦弱無能了吧!但這可是牠的求生之道,有了這個家,至少大半的敵人對牠就莫可奈何,牠只要躲在殼中就沒事了,任敵人在外面叫囂破壞,反正貝殼又不是牠的肉,打也不會痛,而且貝殼也堅固的很。背著這個家,牠可以在很多別的動物無法生存的地方過得很好呢!這麼有趣的小動物,你一定想要更深入的了解牠吧!
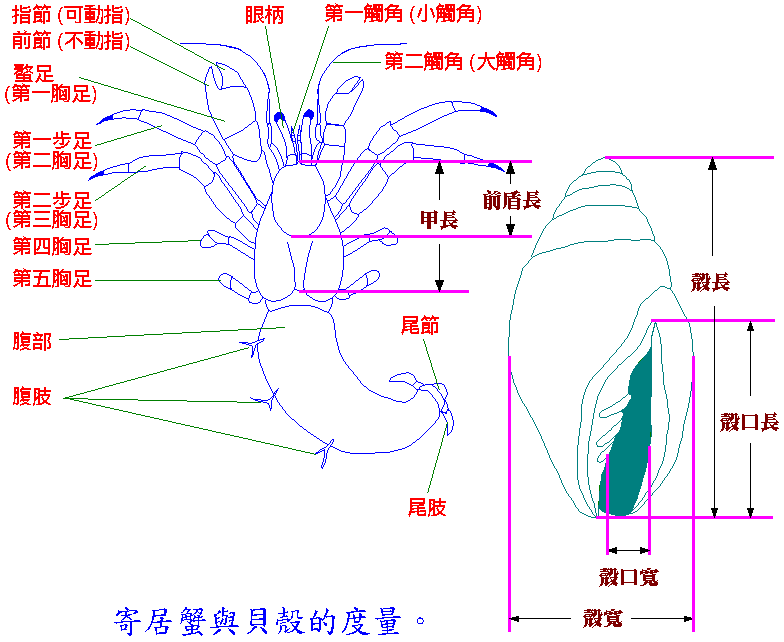
寄居蟹 (hermit crabs) 是一種節肢動物,屬於甲殼類的異尾類,常見的寄居蟹分為三大類:活額寄居蟹科、寄居蟹科和陸寄居蟹科。這三科寄居蟹最大的區別是:陸寄居蟹科的成體是生活在陸地上;而其他兩科都是生活在海中的,這兩科又可以利用左右兩隻螯腳的大小來區分,活額寄居蟹科是左右螯同大或是左螯大於右螯;而寄居蟹科則是右螯大於左螯。墾丁地區出現的寄居蟹除了陸寄居蟹外,其他均屬於活額寄居蟹這一科。基本上所有的寄居蟹都是海生的;像成體的陸寄居蟹雖然是棲息於熱帶海濱附近的陸地上,但仍然會回到海邊釋放幼體 (larvae) 或尋找新殼。全世界大約有 86 屬 800 種的寄居蟹;台灣至 1989 年共記錄有 15 屬 54 種。大多數的寄居蟹是攜帶貝殼移動的,但有少數種類的棲所較為特別,例如 Pylochelidae 科的寄居蟹是居住於某些非貝殼的物體 (包括岩石、竹片、海綿等的穴孔) 中;另外,屬於陸寄居蟹科的椰子蟹 (Birgus latro) 則只有在幼體和前 2 年的小蟹才住在殼中,之後就裸露腹部生活了。寄居蟹最明顯的特徵是腹部不對稱的右旋 (海生的貝殼也是右旋的),腹部柔軟表面沒有甲殼覆蓋 (在貝殼中容易進出) 而只剩下尾部的倒鉤 (尾節和尾肢) (可鉤住殼軸,不被敵人拉出),以及第 4 和第 5 步足的退化 (使身體更能縮入殼內),這使得牠非常適合於背負貝類死後留下的空殼行動。通常從外表是分不出居住在殼中寄居蟹的性別,除非是抱卵的雌性而且卵團露出於殼口外,所以一定要將牠自殼中取出才分得出性別。雌雄寄居蟹最大的分別是雄性生殖孔開口於第 5 對胸足的腰節;而雌性則在第 3 對胸足。
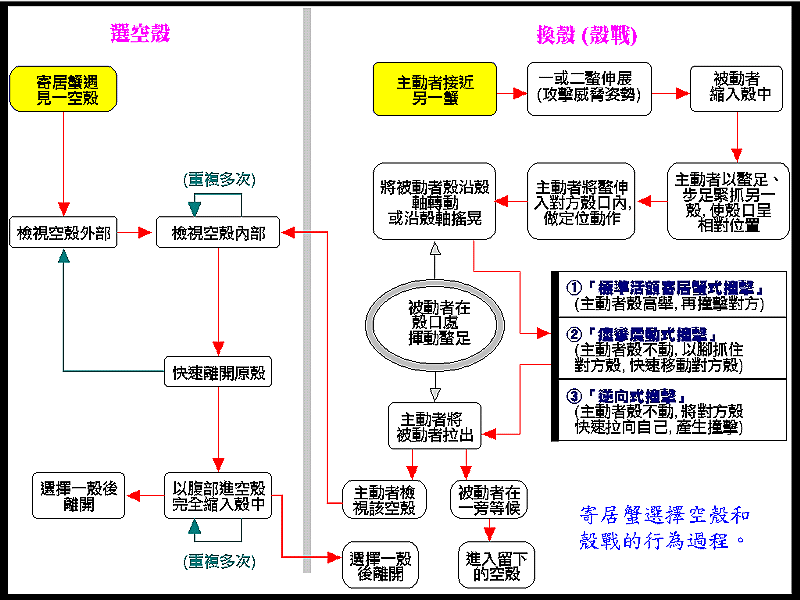
由前述可以知道寄居蟹體型上的改變已使得牠非住在空殼中不可,所以貝類的殼就成為牠必要的資源。大家可能會想到,既然寄居蟹這麼需要貝殼,牠是否會將貝類殺死以取得空殼?但是在自然的情況之下,研究人員並未發現寄居蟹會為了取得空殼而殺死活的貝類,除非是貝類受傷或無法逃走的情形,因此寄居蟹就要靠貝類的自然死亡才能夠得到新的空殼,對於原主人還未死的貝殼,牠可是一點都沒有辦法,頂多在旁邊詛咒牠早點死掉。但是寄居蟹倒是會前往貝類常被捕食的地方,看看能否找到牠理想中的家,尤其是住在貝殼破損得很厲害的寄居蟹更是會前往這種地方,當然,牠被捕食者吃掉的機會也是蠻大的。在大多數寄居蟹可生存的海岸,可供寄居蟹使用的空殼數量都很少,所以可用的空殼對寄居蟹族群的數量是一個限制的因子。能夠遇見貝類剛死亡所留下的空殼的寄居蟹並不多,大多數寄居蟹都要靠與其他寄居蟹 (同種或不同種) 的換殼 (shell exchanging) 才能有新的住家。通常換殼之前會有一連串的儀式化行為 (ritualized behavior),學者們常稱之為殼戰 (shell fighting),其中最特別的行為就是主動的寄居蟹 (通常是較大的蟹) 抓住防禦者的殼口,使兩個殼口面對面,接著主動者利用腹部的力量將自己的殼連續猛擊 (rap) 對方的殼口,想不到吧!寄居蟹竟然是用這種方式來迫使對方讓出殼的。此時防禦者有兩種選擇,一是抵死不從,躲在殼中怎樣也不出來;另一個就是乖乖讓出殼,給主動者挑選。如果是後面的情況,主動者會將對方的殼翻來覆去,很詳細的檢查,甚至會搬過去住看看到底適不適合牠,最後主動者在兩個殼中挑選一個牠最滿意的殼離去,在旁邊觀看的防禦者只好別無選擇的住進剩下的空殼。
一個合適的空殼對寄居蟹是非常的重要,研究人員發現空殼除了保護身體使牠得以成長之外,還能夠提供寄居蟹以下的好處:避免受捕食者所捕食,在地上爬行時能保護柔軟的腹部,避免受溫度變化、缺水、鹽度變化的影響以及保護雌性寄居蟹的卵團。另外,住過小的殼會使蟹的生長受抑制;但是住在過重過大的殼所花費的能量會較多,影響個體的成長,行動也會變得比較笨拙。而當牠身體狀況不同時 (長大了或抱卵時等) 或環境不一樣了,此時,原本合適的家又變得不適當了,因此牠時時要為住處而奔波,這可由寄居蟹一見到貝殼或類似貝殼的物體就立即趨向前去探究一番而略窺牠對殼的迫切需要。
而我之所以會研究寄居蟹也是當初沒想到的。剛進研究所時也如同許多研究生一般,在為尋找論文的題目而煩惱,指導教授莫顯蕎老師提到過寄居蟹的行為生態蠻有趣的,並介紹了些文獻,研讀之後,發現國外的文獻非常豐富,但是台灣只有少量分類上的資料,而寄居蟹與貝殼的關係確實是丁個研究的好題材,由幾次到墾丁浮潛的實地探查,發現實驗的材料不成問題,再加上中山大學海洋科學學院在墾丁青年活動中心內有一棟 “墾丁臨海工作站”,對於實驗的進展有很大的便利,由這些因素才促使我決定踏入此 “異尾類” 的研究領域,成為 “底棲碎食公會 (benthic detritus-feeding guild)” 的一員。
起步總是最難的,首先要面對的就是得挾著剛受完潛水訓練的技術獨自投入大海,在海邊做實驗與休閒玩樂的滋味真是截然不同的,當自己被海浪打得翻來覆去時,就會深深感受到大自然的 “偉大” 力量,人類實在是渺小又脆弱的。我選定位於墾丁國家公園內的 4 個地點採樣,分別是後灣、貓鼻頭、南灣及石牛鼻,從 1988 年 12 月到 1990 年 2 月在這些地點採集寄居蟹及貝類。利用潮水最低的時刻,將採樣用的穿越線 (transect) 綁在潮間帶高處的石頭上,再拉到深約 2、3 公尺的淺亞潮帶,將線綁在珊瑚枝上。採樣的方式是自己位於線的中央,採集左右兩手臂範圍內所有的寄居蟹和貝類。作業時還得時時留心岸上,是否有好奇的小朋友將我紅色的穿越線扯斷 .....。採樣工作最辛苦的地方是在潮間與亞潮帶交界處,有過潛水經驗的人在這個地區上下岸時都會特別的注意,因為這地方的浪最猛烈且周圍銳利的礁石及珊瑚最接近身體,往往一個大浪過來,若不立即警覺的雙手緊抓礁石,是會被無情的浪將身體往前猛推,再用力的往回扯!撞得自己頭昏眼花,甚至傷痕累累。有時因為天氣不佳或低潮時間已接近傍晚,在水中的視野愈來愈模糊,抬頭望望四周,一片混沌,頗有天地初開之氣魄。回工作站後,立刻將樣品放入事先已準備好的新鮮海水中,才鬆了一口氣。
研究寄居蟹的人同時一定得對貝類有所認識,往往為了確定是那種貝殼而翻遍圖鑑,但圖鑑上大多是漂亮完整的標本,自己所採的卻是破損或表面覆蓋有其他的東西,必須花費很多的時間仔細辨認方能確定。接著,你即將會發現寄居蟹的研究怎麼會如此困難!不管你是要鑑定種類、分性別或測量大小,不把牠自殼中取出是不可能辦得到的!你絕對不能用強硬的方式將牠拉出殼,牠的尾鉤會緊緊的鉤住殼軸而使得你得到的東西只是牠的 “前半身”!這當然不是你想要的吧!大部份的樣品我是利用加熱殼頂的方式將蟹趕出,不過得非常的小心,稍一不慎就會使牠們死於非命,所以為了使牠們全身而出,真是絞盡腦汁的和這些小動物鬥智。如果不幸死於殼內,就只好用大型的老虎鉗壓碎貝殼了。將寄居蟹取出來測量的這個階段是很花時間的。由於台灣缺乏有關寄居蟹族群調查的資料,但這卻是除了分類之外最基礎的本土化工作,因此雖然不是論文所必須的資料,我仍對每個樣品做最詳細的記錄,希望能對墾丁地區的所有寄居蟹有最深刻的了解。我分別記錄了寄居蟹和貝殼兩方面的數據,寄居蟹方面的記錄包括了殼適性測試 (詳見後述)、種類、性別、甲長 (是測量沿著甲殼中線的最前緣 (吻端) 到最後緣 (頸縫) 的長度)、前盾長 (只測量甲殼前半部硬質部份的長度)、濕重;而對於貝殼之分類主要是基於貝類的屬,而將貝殼分為若干的殼型 (shell types),另外再記錄殼長、殼寬、殼口長、殼口寬、在海水中的重量、貝殼的狀況 (殼頂破損、殼口破損、殼口有東西堵塞、殼表面有否覆生藻及藤壺...)。
為了解野外的寄居蟹所住的殼是否足以保護其本身,以身體縮入貝殼內的深淺程度來判斷是一種很直接的方式。若寄居蟹無法完全縮入,則寄居蟹可能較容易被捕食或被較大的寄居蟹拖出,而且身體露出的程度愈大,水份的蒸散也愈快。本實驗所判斷的標準是愈能縮入殼內的分數愈低,而露出殼外愈多的分數愈高:
(1) 完全見不到蟹,也就是蟹可完全縮入殼內。
(2)
可見到部份的螯及步足。(3)
可見到整個螯表面,螯表面幾乎與殼口面平行。(4)
螯表面幾乎與殼口面垂直,但螯仍在殼口面之內。(5)
螯突出殼口面。實驗的結果顯示,多數個體 (80%) 的殼適性分數均為 1,也就是墾丁地區的寄居蟹 (以潮間帶和亞潮帶的 C. latens 與亞潮帶的 C. gaimardii 為例) 所住的貝殼多足以保護牠本身,至少在牠完全縮入殼中時,敵人看不見貝殼中有寄居蟹在裡面,就算知道牠在裡頭,要將牠自殼中拉出也不容易。有時可發現殼口破損很厲害的貝殼,據文獻記載,蝦蛄會利用其強有力的攫捕足將貝殼殼口一塊塊敲掉,最後吃掉縮於最內的螺層中的寄居蟹,所以有一個可完全縮入的貝殼,是可以阻止大多數捕食者攻擊的。
分析數據的結果,在寄居蟹的分佈狀況上,共採得 1088 隻的寄居蟹。潮間帶數量最多的是 Calcinus latens (佔 74%);亞潮帶數量最多的是 C. gaimardii (佔 42%),其次是 C. latens (佔 29%)。而在貝類的分佈上,共採得 673 隻貝類,潮間帶和亞潮帶數量最多的同樣是 MOT 型貝類 (此型貝類包括 Morula sp.、Drupella cornus 及 Ergalatax contractus);次多的也都是 Conus。大家可能會問,寄居蟹是不是使用數量最多的那種貝殼?結果並非如此。潮間帶及亞潮帶的 C. latens 都是較常使用 Conus,而較少使用 MOT 型殼;為什麼會這樣呢?
為了解在理想情況之下,寄居蟹會選擇那類牠最喜好的殼,因此進行實驗室的選殼實驗。此部份我以潮間帶的優勢種 C. latens 為材料。自野外採回的寄居蟹利用壓碎殼的方式取出寄居蟹。將裸蟹放入底部舖有小碎珊瑚石的容器中,我是以配對方式 (pairwise) 來置放空殼的,也就是同時放入兩類殼型 (shell types) 讓蟹選擇;若給蟹兩類殼 A、B,蟹喜好 A 殼而不喜好 B 殼;另外再給蟹 B、C 兩殼,若蟹較喜好 B 殼而不喜好 C 殼,則判斷此寄居蟹對此三類殼型喜好的程度為 A > B > C。本實驗置入的殼型是較易採集到而且是蟹較常使用的 (Conus、MOT 型殼、Cypraea、Mitra 及 Drupa)。所置入的殼大小範圍是盡量供給作者所能採集到的殼大小範圍。蟹:殼的數量約為 1:2 至 1:5。給裸蟹一至二天時間選擇。
結果在 Conus 及 MOT 型殼這一組中,有 92% 的個體選擇 MOT 型殼,其他 8% 則選擇 Conus。所以 C. latens 其實是較喜好 MOT 型殼,也就是野外最多產的貝類,但實際上牠在野外並非用 MOT 型殼,是什麼原因呢?由於缺乏實驗的證實,只能根據一些觀察現象來推測:因為 MOT 型貝類的平均體型較小,可能導致較大的寄居蟹無法使用,而且此貝類多生活於潮間帶高處岩礁上,低潮時常曝露於空氣中,顯示牠的生命力很強,而實驗室的觀察也可支持此一說法,同時飼養的 MOT 型殼和 Conus,MOT 型殼能存活很長的時間;而 Conus 很容易就死亡。因此可能是因為 MOT 型殼的大小關係,以及 MOT 型貝類的死亡率較低,才使得大多數的 C. latens 使用次多產的貝殼。
另外我想了解的是寄居蟹是以什麼標準來選擇合適的殼,換句話說,寄居蟹是考慮本身的什麼特徵來選擇具有那些特徵的殼,然後再嘗試以其生態環境的因子加以解釋。蟹-殼的關係是利用統計方法中的複迴歸分析來處理數據,殼方面的介量 (parameters) (也可說是生物上的特徵) 我選擇了殼長、殼寬、殼口長、殼口寬及殼在海水中的重量;蟹方面的介量則有甲長、前盾長及蟹的濕重。由統計分析我可以推測出以下的結論:潮間帶及亞潮帶的 C. latens 是相同的族群。亞潮帶 C. latens 與 C. gaimardii 可能存在有殼的競爭。雖然野外寄居蟹的殼適性分數多為 1,但選殼實驗顯示寄居蟹在有較大的殼可選時,仍會選擇較大的殼,這可能是因為牠選了稍大的殼,牠可以有較長的時間使用這個貝殼,而不必頻頻的更換,萬丁換不到好殼那不是很麻煩嗎?潮間帶 C. latens、亞潮帶 C. latens 與亞潮帶 C. gaimardii 各群體的蟹-殼關係中,與蟹介量最相關的殼介量分別是殼長、殼寬與殼重,也就是這些群體的寄居蟹選殼時,可能會先考慮這些殼的介量。而這些差異可能反應出其生活環境的壓力。潮間帶的寄居蟹由於常會曝露於空氣中,而長螺旋的殼 (與殼長有關) 是會比短螺旋的殼保有較多的水份,由於水的比熱很大,殼內有較多水份的話,當周圍溫度上升時,裡面的溫度就較不會上升得這麼快,因此寄居蟹會選擇一個有最適當殼長的貝殼來當作牠的棲所。而亞潮帶寄居蟹的殼寬和殼重跟殼的重心和殼的厚度有關,可能是亞潮帶的浪流較大,有較重的殼是較不容易被浪流衝走的;而殼的厚度跟是否容易被捕食也有密切的關係,所以亞潮帶的寄居蟹住在有最適當殼重與殼寬的貝殼中是不無道理的。
有關寄居蟹的研究在台灣並不多,已發表的文獻均為分類方面的報告,其他方面的研究則幾乎沒有,但是寄居蟹在各處的海岸都是數量上的優勢種類,而寄居蟹與所住的貝殼有著非常密切的關係,因此是研究生態、行為及演化的適當材料。由於生物間所競爭的棲域 (niche) 通常是很難量化的,將寄居蟹的殼認為是一個微棲所 (microhabitat),用來測試一些生態上的模式 (models) 是很理想的 (殼的各種介量都很容易量化);在行為學上,兩個動物常會因爭奪資源而發生敵對行為,寄居蟹有很複雜的換殼行為 (shell exchanging behavior),也就是所謂的殼戰,兩者所爭奪的資源即是兩方最好的殼資源,但就算是輸者仍有一個勝者認為較不好的殼,因此在行為學上,寄居蟹也是一很好的題材。
研究寄居蟹不到 2 年的時間中,我深深感受到寄居蟹真是一種不簡單的動物,潮間帶的環境那麼惡劣,隨時要曝露於溫度、水份、鹽度等等的危險當中,但就因為牠有了殼,而使得牠能自由自在的存活於潮間帶王國,近代最早有系統研究寄居蟹行為的夏威夷大學學者 Reese 曾說過:殼提供了所有洞穴擁有的好處,但卻完全沒有它的限制。的確,寄居蟹攜帶了一個殼,就如同帶了一個活動的洞穴,不像招潮蟹一類的穴居者只能棲息於特定的地方,而牠卻能利用穴居者所難以使用的環境,若是環境變得太惡劣時,寄居蟹仍可帶著牠的螺殼離開這個環境。因此牠是潮間帶動物中非常成功的一群,也是這些環境中最顯著的成員之一。
最後,希望國內的有志之士多投入台灣本土化的工作行列中。我們只有一個台灣,但是事實上,我們對於台灣的生物並沒有完全的了解,尤其是無脊椎動物,可能連有那些種類都不能確知,更何況進一步的研究。誠然,做分類、生態調查是比較枯燥辛苦的,但是研究過程中仍有不少的樂趣,而且在為以後的研究奠定基礎的考慮之下,這是很值得去做的。想到現在我們的學生還要拿著日本出版的圖鑑來對照它的圖片才能認識我們的生物的話,這不是很令人汗顏嗎?別的不說,就拿寄居蟹的研究來說,我所收集的國外文獻數量已有二、三百篇!研究的範圍從生態、行為、生理等等都有,而且每年均有新的文獻出來,文獻多得真使人看得都快消化不良。因此我希望大家能重視這個問題,使我們的生物學能平均的發展,而不是一窩蜂的往熱門的分子生物走,到頭來可能會失掉我們的根。以保育的立場來看,雖然寄居蟹在海邊的數量很多,但是仍希望大家以欣賞研究的心態來對待這些小動物,不要任意的採集傷害牠們。尤其近年來,因為熱帶海岸樹林的砍伐、人類的侵入以及海邊的污染等等,使陸寄居蟹的棲息地受到很大的破壞,數量也急劇的減少,但是我們卻常可在路邊攤上看到販賣的陸寄居蟹。而另一種椰子蟹,有人認為牠們的精巢卵巢具有滋補的效果,大量捕捉的結果,現在台灣島上大概沒有了,而蘭嶼、綠島等離島的數量也變得稀少,所捉到的個體也愈來愈小,為了老饕的口腹之慾,甚至有人從南洋群島走私進口,你說這是不是人類的悲哀!
BOX
寄居蟹在分類學上的地位:
PHYLUM Arthropoda 節肢動物門
SUPERCLASS Crustacea
甲殼上綱CLASS Malacostraca
軟甲綱ORDER Decapoda
十足目SUBORDER Pleocyemata
抱卵亞目INFRAORDER Anomura
異尾下目FAMILY Diogenidae
活額寄居蟹科FAMILY Paguridae
寄居蟹科FAMILY Coenobitidae 陸寄居蟹科
BOX
墾丁珊瑚礁地區常見的寄居蟹:(讀者若欲進一步認識這些種類,可以參考日人三宅貞祥 1982 年所著的 “原色日本大型甲殼類圖鑑 (I)”,通常珊瑚礁所產的寄居蟹依外表的顏色即可分出種類。)
Calcinus 屬
此屬的寄居蟹分佈很廣,從潮間帶高處到淺亞潮帶均可發現。大多數的種類是較不活躍的,常會倒吊於岩石下,或在陡峭的斜坡上。在形態上,屬於 Calcinus 的種類都非常相似,有的要靠不同的顏色型式來區分。此屬的特徵是顏色鮮豔美麗,因多出現於珊瑚礁地區,所以日人稱此屬寄居蟹為珊瑚寄居蟹。眼柄細長,左螯表面平滑,且大於右螯。兩螯的指尖是磁質的 (porcellanous)。
身體大致為橄欖綠色,步足指節前半白色、後半黑色,眼柄粉紅,角膜 (眼柄最前端) 部份為黑底綴有白點。在潮間帶及淺亞潮帶的數量均非常豐富,潮間帶以高潮間帶的岩石下及岩礁的凹窪處有較高密度的族群;淺亞潮帶則多棲息於珊瑚叢枝間。
各胸足深棕色,背甲淡橄欖綠,眼柄後半深棕、前半寶藍,角膜部份為黑底綴有白點。在淺亞潮帶數量最多,多棲息於珊瑚叢枝間及岩礁上。低潮間帶偶而也可找到單獨活動且較小的個體。
全身白色,綴有橘紅色點,步足指節亦為橘紅,眼柄淡粉紅,角膜部份黑底綴有白點。數量不多,發現於淺亞潮帶的珊瑚叢枝間。
在淺亞潮帶的珊瑚叢枝間,常可同時發現 C. latens,C. gaimardii 及 C. minutus,所居住的貝殼表面常有紫紅色的珊瑚藻 (coralline algae) 覆蓋著,有時此種藻類長得過厚甚至無法分辨出貝殼的形狀,此種貝殼與周遭環境的顏色非常相近,可以提供寄居蟹良好的隱蔽色。另外在珊瑚叢枝間的寄居蟹有一躲避敵人的行為,當有干擾時,牠會立即縮入殼中,使自己掉落到珊瑚叢枝中央,讓敵人很難捉到牠。
各胸足深棕色、尖端為白色,左螯巨大,指節前半部的白色尤為明顯,背甲淡橄欖綠,眼柄基部橘色、後 1/3 淡藍、前 1/3 橘色,角膜淡藍。只出現於高潮間帶的碎石上及岩礁凹窪處,常會離水而曝露於空氣中,由於左螯非常巨大,通常住在殼口寬大的貝殼中。高潮間帶岩礁上數量最多的是蜑螺 (Nerita sp.),其體型小,易於背負,同時殼口寬大,所以成為此種寄居蟹最常居住的貝殼。
Clibanarius
屬此屬的寄居蟹只棲息於潮間帶。顏色多晦暗。特徵是兩螯細小,表面有小瘤,有蹄形的角質化 (corneous) 指尖。
各胸足為深綠色,步足指節為黃色,出現於高潮間帶,數量比 C. humis 稍少。
與前種的形態非常類似,但胸足為深褐色,步足指節為橙色。個體小,在高潮間帶岩石下常有很高的族群密度。
在採樣時,常可同時見到 Calcinus latens,C. laevimanus,Clibanarius virescens 及 Cl. humis 4 種寄居蟹,這些種類在低潮時,常會曝露於空氣中。
是珊瑚礁可發現最大型的 Clibanarius 寄居蟹。胸足和甲殼為深紅褐色,長有短粗毛,眼柄橘色。數量不多,常單獨出現於高潮間帶岩礁上,也會在水邊行動。
眼柄和甲殼有黃、褐縱向條紋相間。多發現於潮間帶岩礁凹窪處,頭胸甲非常扁平,多住於殼口狹窄的貝殼中,如芋螺 (Conus sp.) 及鳳凰螺 (Strombus sp.)。由於多棲息於靜止的水中,貝殼表面常有污物附著。
Aniculus aniculus
大型個體,左右螯腳同大。全身大致為橄欖色,螯腳及步足有橫向的紅色條紋,全身長有長剛毛。常可在 3 至 5 公尺深的亞潮帶發現單獨行動的個體。多住於大型、厚重的蠑螺 (Turbo sp.) 中。
Dardanus
屬此屬為大型個體,身體長有剛毛,左螯大於右螯,通常單獨出現並在珊瑚礁的特定區域遊蕩。一水族箱中若有兩隻以上的個體,常會自相殘殺,最後只剩下一隻。
全身橘紅色,綴有許多黑緣的小白點,眼柄深橘紅色。在潮間帶可發現。
胸足紅色雜有小白點。出現於潮間帶,特徵是螯腳與胸足的腕節無毛部份有鮮明的藍色斑點。身體非常扁平,多住於殼口狹窄的貝殼中,如芋螺及寶螺 (Cypraea sp.)。
另外也可在潮間帶沙底處找到殼上附生有海葵的 Dardanus sp.,若牠被干擾時,會以非常快的速度逃跑,殼上的海葵也會放射出長絲狀的刺絲胞以幫助抵禦捕食者。當寄居蟹要換殼時,牠會將原來殼上的海葵移植到新殼上,擁有海葵的寄居蟹較不易被章魚所捕食,同時也較不會受其他怕海葵螫的寄居蟹侵擾。
在海濱高潮線以上的砂礫地及樹叢間常可找到為數眾多的陸寄居蟹,牠的左螯巨大,無法如其他寄居蟹般的完全縮入殼中,但遇危險或乾旱時可當作類似貝類的口蓋,有效的保護自己及防止水份的喪失。
●照片
(1,2)
墾丁的海岸使人有想投入的衝動。(3)
水上及水下的世界是截然不同的。(4)
潮池中的生態條件比較溫和,不會受到波浪的衝擊,低潮時也能保有水份(5)
潮池的水溫變化極大,而寄居蟹對溫度的忍耐力也極強,曾有人做過實驗,C. latens 的致死溫度是 38-40 ℃,但當寄居蟹覺得受不了時,就早已背負著螺殼離開這個不適宜的環境了。(6,7)
潮池是充滿生機的地方,豐富的藻類,提供大量潮間帶動物基本的食物。(8)
實驗地點之一:後灣海岸。(9)
實驗地點之一:貓鼻頭潮池。(10)
實驗地點之一:南灣海岸。(11)
實驗地點之一:石牛鼻海岸。(12)
實驗材料之一的 Calcinus latens,是墾丁地區數量的多的寄居蟹。(13)
實驗材料之一的 Calcinus gaimardii 是墾丁地區淺亞潮帶數量最多的寄居蟹。(14,15)
海濱的礁石凹窪處常有大量的寄居蟹聚集。(16,17)
潮間帶常見的菊石珊瑚 (Favia sp.),在退潮後常曝露於空氣中,C. latens 常會聚集在死亡的珊瑚頭 (coral heads) 上。(18)
基本上寄居蟹是食碎屑的,任何食物都吃。(19)
墾丁地區 C. latens 最常使用的是芋螺屬 (Conus sp.) 的殼。(20) Dardanus megistos
所蛻下的甲殼,圖中所斷的左螯會在數次蛻皮後恢復原大小。(21)
眼部及額部。(22)
右螯。(23,
24) 沿岸底拖上來的寄居蟹 Diogenes spinifrons,由其羽狀的第二觸角鞭可推知牠是濾食性的動物 (濾食懸浮於水中的顆粒) 。(25)
小心!我可是左撇子的重量級拳王 - Calcinus laevimanus,在潮間帶高處的岩石下常可發現。(26)
這個家不錯吧!(27)
瞧!我像不像戴了一頂大草帽 - C. gaimardii。(28)
喘死了!背著房子爬山有夠累 - Cl. virescens。(29)
上山難,下山更難!稍不留神可會一路滾到底。(
附註)有關評論寄居蟹行為生態方面的文獻有:
- an introduction to the natural history of hermit crabs. Field
Studies 7: 189- 238.
寄居蟹的形態圖。
以歐洲水域最多產的寄居蟹 Pagurus bernhardus 為例 (據 Lancaster 1988)。此種寄居蟹屬於寄居蟹科的一種,注意牠的右螯大於左螯。
Utilization of Shell Resource by Hermit Crabs in Kenting, Taiwan
(Hsi-Te Shih)
Two species of hermit crabs, Calcinus latens and C. gaimardii, were common in Ken-Ting shore, southern Taiwan. In number, the former was dominant at intertidal area, but was subordinate at subtidal area. The latter was dominant at subtidal area. The shell resource utilization by these species in natural environment was studied and compared with result for laboratory shell selection experiment of the intertidal C. latens. Shell parameters included shell length, shell width, aperture length, aperture width, and weight in seawater. Crab parameters included carapace length, anterior shield length, and wet weight. Multivariate analysis was applied to describe the relationship between crab and shell parameters.
By statistical analysis, intertidal and subtidal groups of C. latens were the same population and there was competition between subtidal groups of C. latens and C. gaimardii. The result of shell adequacy test showed that most crabs lived in adequate shell at least by internal volume. However, shell selection experiment proved that crabs would select larger shells if they were available. The most correlative shell parameter of intertidal C. latens, subtidal C. latens, and subtidal C. gaimardii was shell length, shell width, and shell weight, respectively. These results may be responded to their environmental stress.
n
related topic: Utilization of shell resources by the hermit crabs, Calcinus latens and Calcinus gaimardii, at Kenting, southern Taiwan| 寄居蟹資訊網 | 台灣博物學家 |
Copyright © 2011 Hsi-Te Shih
{kind=link}
{kind=link}
{kind=link}